www.koi-unleashed.co.uk
Koi Physiology
By Spike Cover
Introduction
A short chapter on koi physiology can only begin to cover the subject. With this in mind, we will address primarily those things that are most directly relevant to the goals of the hobbyist,
The Cardiovascular System
The function of the circulatory system is to deliver materials to and from the cells. The system has four major elements: the vessels, the blood, the heart and the control system.
Vessels - In general, fish do not have a separate respiratory circulation,
as is found in mammals. Blood flows
from the to the gills and then is distributed to the head and body. Normally,
blood is distributed from arteries to arterioles to capillaries, and then is
collected from the capillaries to venules to veins. However, blood from the intestines is distributed to the liver
where it again passes through capillaries prior to returning to the heart. This passage through a second set of
capillaries on the way back to the heart is called a portal system. The portal
system of the liver is present in most species, including humans. The liver or hepatic portal system plays a
role in digestion and in removal of toxins from the blood. However, unlike
humans, fish and other cold-blooded species also have a renal (kidney) portal
system in which the blood from the caudal (rear) and sides of the body is
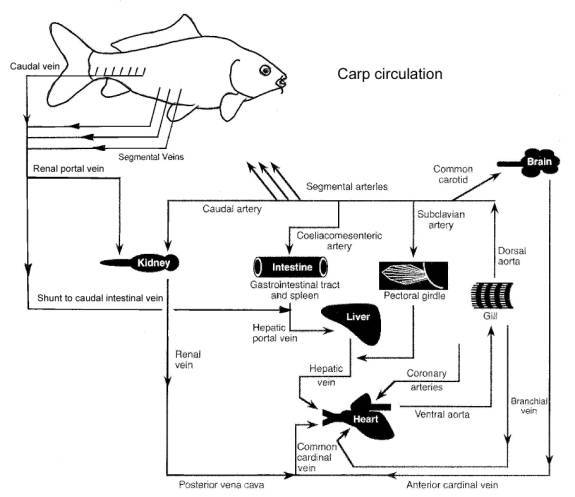
filtered through the kidney prior to returning to the heart. In the carp (koi), a portion of the caudal
vein venous blood flows through the kidney via the renal portal vein while some
is shunted to the caudal intestinal vein and subsequently through the liver The
diagram below is representative of the koi circulation system and is reproduced
here with the kind permission of Koi Carp magazine.
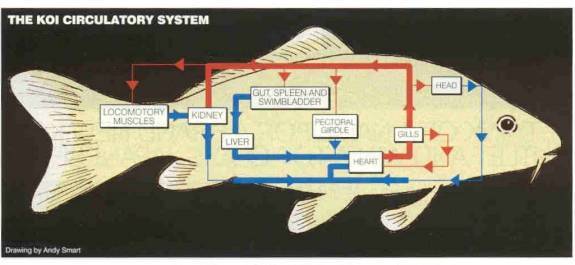
The diagram below is another way to represent the same blood flow system and paths within the koi.
Blood - The hematocrit of blood is the percentage of the total blood volume
occupied by the red blood cells (RBCs) and is typically found by centrifuging
blood in a capillary tube forcing RBCs to the bottom of the tube.
The ratio of the length of the RBCs’ volume to the total length of
the blood in the tube is taken and represents the hematocrit (or packed-cell
volume, PCV). The blood of common carp (koi) has a hematocrit
of about 32-35%. A thin whitish layer
will be formed direct on top of the RBCs and is referred to as the ‘buffy
coat’ and is made up of the white cells within the blood. The transparent fluid above the buffy coat
in the tube is referred to as the supernatant or plasma. If plasma has its clotting factors removed,
the remaining fluid is referred to as serum.
The hemoglobin content of carp blood is about
8.2 g/dl. Diseases can change this and other hematology
numbers. The salinity of the blood is 0.9%
Heart - A koi's heart is a two-chamber organ having a ventricle chamber and an atrium chamber. The extra two chambers that precede and follow the two pumping chambers are called the sinus venous and the bulbus arteriosus. They are smaller than the pumping chambers and function as compliant accumulators (elastic and expansive when pressurized, i.e., balloon-like) to smooth out pressure surges and protect the cardiovascular system. They are not intended for pumping blood as they have non-muscled walls.
Control System - Control of the cardiovascular system includes local and remote functions. However, these systems are complex and beyond the scope of this course. Suffice to say it is the blood pressure that is regulated in the short term by the remote
System and since most of the organs are in parallel, localized control of blood flow through any given organ is in response to the organs needs.
Gas Exchange
A portion of the following is reproduced with permission of Paul Maslin’s from his web page at: http://www.csuchico.edu/~pmaslin/ichthy/fshrsp.html
Respiration takes place at the gills via diffusion. Diffusion exchangers provide a passive exchange, operating on physical laws.
![]()
R - Rate of diffusion.
D - Diffusion constant (a function of the material the substance is diffusing through.)
A - Area of exchange
![]() - Difference in partial pressure between the
two solutions.
- Difference in partial pressure between the
two solutions.
d - distance involved.
Of these, A, ![]() ,
and d can be most easily modified to control the rate of diffusion.
,
and d can be most easily modified to control the rate of diffusion.
Area
The larger the surface area, the more rapid the diffusion. In general the gill area is proportional to the amount of red muscle, more active fish having a proportionately greater area. A mackerel has 1000 sq mm /g body weight; a toadfish has 57 sq mm/g body weight. The problem with increasing the area is that water also diffuses and the amount of work the kidney has to do for osmotic regulation increases proportionately to the gill area.
Distance
Fish have reduced this to a minimum. One layer of epithelium; one layer of endothelium.
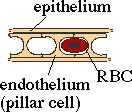
The secondary lamellae (thin, leaf-like protrusions from the gill filament) are best described as a sheet of blood separated by an array of posts (the pillar cells) which keep the lamellum from ballooning, maintaining it one red blood cell thick. Only about 2 μm of tissue thickness separates the water from the blood stream. Any increase in this distance will decrease oxygenation, e.g., gill tissue hyperplasia (thickening of the gill tissue) or increase mucous coating. Because this tissue thickness is so slight, often koi under great stress, especially from over handling, will seep blood from their gills.
![]()
Fish as a group have done a great deal to maximize ![]() .
The first step is to keep both blood and water moving. If either stopped,
it would quickly reach an equilibrium with the other and diffusion would go
to zero.
.
The first step is to keep both blood and water moving. If either stopped,
it would quickly reach an equilibrium with the other and diffusion would go
to zero.
1. The blood pump: the heart pumps in a discontinuous fashion, squeezing out a spurt, then having to relax and refill before squeezing out the next spurt. However the bulbus arteriosus is made of elastic fibers and expands to receive the entire output of the ventricle. It's elasticity keeps pressure on that blood and keeps it flowing smoothly while the ventricle refills (minimizing pressure fluctuations in the system).
2. The Water Pump Water also moves
continually across the diffusion surface. There are two pumps, a buccal force
pump and an opercular suction pump and two one-way valves, an oral valve just
inside the lips and an opercular valve reinforced by the branchiostegal rays.
The buccal pump consists of the hyoid apparatus, which can be depressed to draw
water into the mouth, then raised to force it out. However as the water starts
to exit the mouth, it inflates the cusps of the oral valve, popping it closed.
The water then can only exit by way of the gill slits to the opercular cavity.
As the opercals are flared out, the opercular valve is pressed against the
body, preventing the entry of water so that water must enter from the buccal
cavity across the gills. With the two pumps operating slightly out of phase a
constant difference in pressure can be maintained in the two chambers resulting
in a constant flow of water across the gills.
In other words, water flows into the mouth, over the gills and out through the opercula. The one-way flow is more efficient in providing oxygen to the fish. Ventilation rate can be determined by observing opercular movements. Since the concentration of carbon dioxide and related substances (carbonic acid/ bicarbonate/ carbonate) are very high in water, fish tend not to be responsive to changes in carbon dioxide, but are very sensitive to changes in oxygen. Low oxygen levels will cause a noticeable increase in ventilation rate, which can easily be observed. Other causes can include damage to the gills, exercise or fright.
![]()
A resting teleost fish typically removes 80% of the oxygen from the
water passing over its gills. Some experimental results for the tench showed a
mean efficiency of 51% oxygen removal. Reversal of the water flow reduced the
efficiency to 9%. A typical mammal,
such as a human removes only 10 to 20 % of the oxygen from air. Why such a
contrast in efficiency? Consider the medium.
Water is 800 times as dense as air. Air normally contains 20% oxygen (200,000
ppm). Water, by contrast contains only about 10-ppm oxygen and under stagnant
conditions this may drop much lower. Fish have to be more efficient.
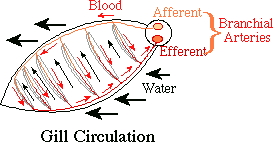
Also, in order to conserve energy when the complete gill function is not needed, koi take advantage of basal channels in the gills that allows a portion of the blood (up to about 40% when the fish is at rest) to bypass the high-pressure drop of the gills and thus requires less energy to perfuse the body. This basal channel is a shunt around the gills and is shown in the figure below. When needed, the entire gill can be utilized to supply oxygen to the fish. Increasing the number of perfused lamellae by shutting off blood flow through the basal channel is called “lamellar recruitment.”
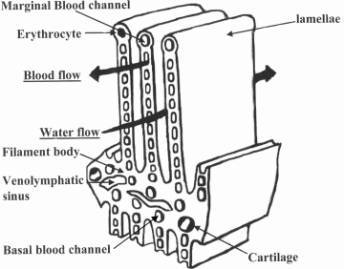
If fish gills are so efficient and air contains so much more oxygen,
why do fish suffocate out of water? Surface tension of water clinging to the
gill collapses the lamellae and the effective surface area is reduced to a tiny
fraction of normal. Catfish and ells survive better than most other fish
because they have cartilage supports for the primary filaments and have the
secondary lamellae thickened and separated more than usual.
4. Respiratory Pigment Oxygen diffuses
into the blood according to the difference in dissolved oxygen (DO) between
water and blood. Until it is saturated,
hemoglobin in the blood will react with the oxygen to form oxyhemoglobin,
tending to keep the DO low and ![]() high.
This is because O2 bound to hemoglobin is no longer in solution
and does not affect its partial pressure.
As a result, one ml of blood will carry as much oxygen as 15 to 25
ml of water. The reaction is reversible; as oxygen diffuses out of the blood
into the tissues, more is released from the oxyhemoglobin.
high.
This is because O2 bound to hemoglobin is no longer in solution
and does not affect its partial pressure.
As a result, one ml of blood will carry as much oxygen as 15 to 25
ml of water. The reaction is reversible; as oxygen diffuses out of the blood
into the tissues, more is released from the oxyhemoglobin.
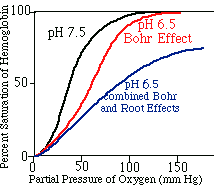
Hemoglobin exhibiting the Bohr effect will require a higher oxygen tension to fully load under conditions of low pH. Hemoglobin showing the Root effect can never be fully saturated with oxygen when CO2 is high.
Normal tissues (muscles, etc.) are high
in carbon dioxide and low in oxygen.
Gills are normally low in carbon dioxide
and high in oxygen.
Hemoglobin loads on the high curve and
unloads on the low curve.
Fishes
adapted to slow water habitats, such as carp where low oxygen and high carbon
dioxide content are normal, have blood with weak Root/Bohr effects.
An excellent
explanation of these principals and effects may also be found on the web site of Dr. Richard Strange at: http://web.utk.edu/~rstrange/wfs550/html-con-pages/I-oxygen-in-blood.html
Osmotic and Ionic Regulation
To understand the challenges faced by our koi (and other freshwater fish), we need to understand a bit about diffusion and osmosis. Diffusion is the movement of molecules from a higher concentration to a lower concentration. Osmosis is the passing of a solvent through a semi-permeable membrane from a higher concentration of the solvent to a lower concentration of the solvent. In our case, the solvent is water. The pressure required on the ‘migrating to’ side of the semi-permeable membrane to just stop this movement of water is called the osmotic pressure. If the membrane is also somewhat permeable to certain dissolved substances (like most living tissues/membranes are), water will move in one direction and the dissolved substance will move in the other. These processes are referred to as passive and do not require the input of energy.
Koi like all freshwater fish, have body fluids that are hypertonic to their environment (i.e., they have a higher salt content in their body than the water in which they normally live).
The two major osmotic and electrolyte problems in freshwater fish are the elimination of excess water and maintaining the proper salt concentrations within their bodies. This process is sometimes termed “osmoregulation.” Excess water is expelled in the form of urine. Freshwater fish may produce more than 20% of their body weight in urine each day. This compares to terrestrial vertebrates that produce up to 1.5% of their body weight per day as urine. The urine of freshwater fish is very dilute being primarily water with much of the dissolved materials conserved by reabsorbtion back into the fish’s blood prior to leaving the caudal kidney.
Unless damaged, much of the koi’s surface is relatively watertight. The majority of the ‘leakage’ occurs thru the gills and the surfaces of the internal cavities that have contact with water, e.g., the intestines. Most of the water that is taken across the gills is done so in conjunction with the exchange or transport of ions. There seems to be some disagreement among experts on how it happens but what does seem to be agreed upon is that for osmoregulation in freshwater fish, sodium and chloride ions plus water pass into the blood through the gills and hydrogen and bicarbonate ions (carrying the excess CO2) are excreted into the surrounding water.
Since koi are hypertonic as compared to their normal environment, maintaining osmoregulation requires energy (in the form of adenosine triphosphate or ATP ‘pumps’). A large amount of the energy used for basic metabolic chores (standard metabolic energy) is used for osmoregulation. This means that even when a koi is resting, it is using energy to maintain its water and salt balance (along with other functions like respiration and digestion). Consequently, anything that increases the demand for osmoregulation, can be detrimental to the fish, especially if it has other problems, e.g., is under attack by pathogens. It is also easy to see why distilled water that contains no salt would kill a fish due to a lack of ions, which would preclude osmotic regulation. Conversely, it’s relatively easy to see how adding salt to the water can ease the energy demand on a sick fish thus allowing the energy conserved to be expended fighting pathogens and repairing damage.
Any damage to the environmental barriers or to its hemeostatic (maintenance of normal physiology) mechanisms tends to result in water accumulation that causes tissue edema and swelling.
Digestive Function
The food starts through the digestive system by passing thru the pharyngeal teeth at the back of the throat (on the bottom) and that can grind or break it up as need be against a hard cartilaginous pad (on top) at the base of the skull. The food needs to be ground so that the digestive juices can more easily process it. From time to time new teeth grow to replace the older ones.
Koi have no stomach but rather a long intestine that extends from the end of a short, wide esophagus to the anus (vent) and is about three times the length of the fish in adult koi. This allows them to extract much of the nutrients from the food by the time it exits the fish and to process a variety of materials that, in nature, includes a lot of plant material. By comparison, the length of the intestine in very young fish is only about the length of the fish’s body and is therefore better suited to the digestion of more protein-rich food.
The intestine is lined with tiny finger-like projections that increase the surface area for greater absorption of the nutrients. The intestinal wall also contains mucus-secreting cells that help lubricate the food and protect the intestinal wall from the harshness of some of the material ingested. Even so, the cells lining the intestines are replaced every few days because of damage by the food passing through the gut. The entrance to the intestine contains receptors that, when stretched, trigger a kind of a wave motion in the intestine to move the material along. This motion is called peristalsis.
Digestive juices (enzymes) are produced primarily by the pancreas, which is located as cellular islands within the liver and introduced into the gut thru a duct. Other enzymes are produced in the lining of the intestine. Also naturally occurring flora of bacteria in the gut help to digest and absorb the nutrients from the food. The possible negative impact of antibiotics in the feed on this bacterial flora should be taken in consideration when formulating treatment plans and in particular when contemplating using medicated food.
A common misconception is that, during cold weather, food will sit in the koi gut, putrefy and cause death. Koi have different groups of enzymes called isoenzymes that can digest food over a wide range of temperatures. Koi, like other poikilotherms (cold blooded animals) do not need as many calories at cold temperatures. Their metabolism is slower, so bodily functions slow down considerably. While digestion does occur, it occurs at a much slower speed. This means that the food will be in the intestine longer, excretion is delayed, and little or no resorption of eggs will likely occur (see the next sentence for potential ramifications). The reasons for not feeding koi (fasting them) at colder water temperatures are controversial but some of the opinions are: a) it is difficult to judge how much the fish need and it’s easy to overfeed and cause water quality problems especially since the filters are relatively inactive in cold water; b) fasting allows fish to use up the possibly unhealthy fat put on over the summer; and c) females tend to resorb eggs during fasting. Breeders tend to agree that fasting is valuable.
Nitrogen excretion
Because ammonia is a potent neurotoxin, it cannot be allowed to accumulate to any substantial levels in the fish. For most fish, ammonia excretion is easily accomplished by direct diffusion from the plasma across the gill tissue to the large sink of the surrounding water. This diffusion is probably enhanced by the acidification of the gill boundary layer that acts to convert NH3 to NH4+ (this process appears coupled to CO2 excretion). Even though fish are ammonetelic, that is they excrete most (up to 85% or 90%) of their waste nitrogen as ammonia, the exact mechanisms by which ammonia exits the fish are still not completely agreed upon by experts. While the mechanisms are not entirely clear, what is clear is that the gills are the major excretory organs for ammonia and about 70% of the ammonia given off by Teleost fish is done so through the gills.
Kidneys
The anterior, cranial or head kidney is an immunologic organ, producing cells and substances that fight disease.
As mentioned prior, the (caudal) kidneys play an important role in osmoregulation by excreting excess water as dilute urine and reabsorbing needed salts back into the blood prior to excretion. In freshwater fish, control of blood pressure and flow are major determinants of urine flow.
Since the kidneys are excellent filters, they are often used to detect toxins and bacteria. Bacterial cultures of the kidneys in euthanized fish have been the standard procedure.
Liver
Nutrients broken down and absorbed through the intestinal wall are transported to the liver by the blood that picks up the nutrients in via the network of tiny capillaries that surround the gut.
The liver has many functions. It acts as a reservoir for storing digested food as glycogen that can be further broken down into glucose. Glucose can be used for energy. Some of the food delivered to the liver is further processed and broken down into smaller units. Ammonia is produced from the breakdown of proteins into amino acids which are further broken down to ammonia and, to a lesser extent, urea. This is one reason to avoid overfeeding your fish in cold water, i.e., to avoid excessive ammonia production when the filters are not functioning well. By contrast, overfeeding when the water is very warm may also cause problems but due to a lack of oxygen. Further, since lipids and fats are also stored in the liver, excessive feeding can lead to a condition known as ‘fatty liver syndrome.”
The liver cleans up the blood by breaking down old blood cells and detoxifying substances it is able to modify. Toxins which it can’t breakdown are sometimes stored in the liver. This is why toxic metals tend to accumulate in the liver and lead to degeneration and/or disease of this organ. Therefore, removal of heavy metals from inlet water or the addition of substances to detoxify them can promote liver health as can controlling the diet to prevent fat build up in the liver.
The liver also produces bile that is necessary for proper digestion. Bile is made by breaking down various substances one of the most significant of which is the hemoglobin from spent red blood cells. The greenish-yellow color of bile is derived from the breakdown of hemoglobin.
Cholesterol, which plays a major roll in the stabilization of nerves and cell membranes, is produced in the liver.
The liver metabolizes or breaks down hormones, the chemical messengers released into the blood. Once these messages have been delivered, the signal needs to cease or the targeted organ(s) will continue to react.
Swim bladder
The swim bladder is the major organ of buoyancy, but it also plays a role in sound reception. The swim bladder is comprised of two chambers, the caudal (rear) and cranial (forward) chambers, connected by a small duct. The cranial chamber is elastic and is primarily responsible for allowing the fish to move vertically in the water while the caudal chamber remains at a constant pressure to maintain a buoyant level. Koi are able to add or release gas from the swim bladder through the pneumatic duct, a connection from the gut to the swim bladder, and consequently may over-inflate the bladder if they gulp air due to an oxygen deficiency. Over-inflation will cause the fish to remain at or near the surface; under-inflation will cause the fish to sink and require swimming efforts to rise. Due to the high amount of connective tissue, repair after damage to the swim bladder is often slow or incomplete.
Hematopoietic System
Koi have no bone marrow but there is a hematopoietic system, which produces both red and white blood cells. The organs involved include the spleen, the kidney interstitium (surrounding tissue), the head kidney and the thymus. Red blood cells are produced in the interstitium of the caudal kidney, the head kidney and the spleen. Fish lack a true lymphatic system but do have lymphocytes, both large and small. These cells are produced in the thymus, spleen and head kidney.
Immunity & Defenses
All animals, including fish, share their environment with numerous parasites and other disease-causing organisms. Parasites are organisms that live off a host and provide no benefit to the host (although some hold that stimulating the immune system is a benefit). It is their goal to invade the host, develop and reproduce. It is the goal of the host to prevent this or minimize the damage. Pathogenic (disease causing) organisms range in complexity from simple viruses through a range of bacteria up to the much more complex arthropods. Fish have highly developed defenses against disease organisms.
This sub-section deals with the immune systems in koi and the involuntary defense mechanisms they employ to cope with the continual challenge from pathogens. The defense factors can be separated into two systems, innate and acquired.
Innate Defenses
The innate factors are relatively older from an evolutionary standpoint and are essentially constant, i.e., they do not change in nature or intensity based on whether, or how many times, a parasite has come in contact with the fish. Externally, the first line of defense are barriers to entry and are composed primarily of skin, scales and the mucous membrane (also called the cuticle). Internally, the barriers to entry are the mucosa (epithelial covering of internal cavities).
Should a pathogen penetrate the external barriers and enter the body, a second group of defense factors take over. Macrophage-like cells are large phagocytic cells (from the Greek phagos meaning “to eat”) that roam the blood, tissue and organs of the fish and clean up damaged host cells and foreign invaders by engulfing them and delivering lysing enzymes to breakdown the damaged cell or invader. Also a number of proteins circulating in the blood serum (the non-cellular portion of the blood) are known as “complement” because they normally complement the cellular defense in response to an antigen-antibody coupling in a cascading sequence known as the “classical” pathway. However, the complement system can react without the antibody in a sequence known as the “alternative” pathway. Both pathways end in creation of a unit known as the membrane attack complex. This complex inserts into the wall of the target cell and creates a pour or passage thru which fluids and other substances can flow. As a result, the cell inflates and bursts. This system is useful as a defense against invading bacteria, particularly types not previously experienced by the fish. The products of the complement activation reactions also cause the inflammatory response (redness and swelling).
Acquired defenses
Acquired defenses are those that the fish develops as a result of a threat (almost any foreign body or substance) entering the body. While a complex series of things occur within the fish in preparation to defend itself, it is sufficient to say here that cells and substances within the blood ‘learn’ to recognize the particular invader, develop defensive measures and are better prepared to cope with it on a second appearance of the same invader.
This acquired or ‘learned’ immunity is the basis of vaccinations where a portion of a pathogen, or a complete (dead) pathogen, is presented to the system that, in response, then develops specific defenses against that agent.
Effect of Temperature on the Immune System
Koi
are very susceptible to disease between the temperatures of 47º F and 62º
F. Below 47º F the immune system is not fully
functional but the pathogens are relatively inactive. Above 62º F, the immune system is functional, and can normally reduce
the pathogens. The critical range,
also known as “Aeromonas alley,” represent the range over which the pathogens
are active but the koi’s immune system is compromised. Koi with a compromised
immune system, regardless of cause, will be less able to adequately fight
disease.
Stress Response
Stress in fish is similar to stress in people. The responses to stress are part series of physiological reactions called the general adaptive syndrome. This syndrome is divided into three phases: the alarm reaction when stress hormones are released; a stage of resistance during which adaptation occurs; and if the animal cannot adapt, a stage of exhaustion followed by death. These phases were first observed in terrestrial animals and similar, but not identical however, reactions have been observed in fish.
Fight or flight
When a fish is being pursued, the initial stress response in almost all instance results in an immediate increases the available energy, which may allow it to escape. As a fish is stressed and hormones are released, various effects take place. Some of the more obvious effects are on the circulatory system with an increase in cardiac output and a constriction or dilation of the blood vessels. During periods of stress, one of the main physiological effects is to increase blood flow to specific organs. This results in a greater uptake of oxygen (remember lamellar recruitment?) therefore making more oxygen available for the metabolic demands associated with the stress.
Hormones (catecholamines) also influence the metabolic breakdown of glycogen to glucose in the muscle and liver. In the muscle, this effect produces and almost instantaneous source of energy. However, the same hormone also appears to increase the permeability of the gills to water and ions causing the fish to take on more water and loose ions. The release of other hormones (corticosteroids) also affects both osmoregulation and metabolism. The biggest effect on metabolism is the breakdown of glycogen to glucose and the breakdown of proteins; again providing quick energy.
Chronic effects
The stress in most acute situation is short term. The fish either gets away or is caught. If it escapes, it can recover. Fish in captivity, however, are often subjected to long periods of stress. Capture, crowding and changes in water quality have all been shown to produce stress responses in captive fish. All these situations can lead to osmotic imbalance (osmotic shock). A fish can recover from (correct) the effects of short-term osmotic imbalance but may die if the stressors persist. Stress often suppresses various aspects of the immune system such at phagocytoses and intracellular killing by macrophages. Frequently an outbreak of disease occurs after moving fish from one environment to another.
Fish acclimated to capture can show a reduced mortality upon being moved. However too frequent capturing can lead to problems.
Fish exposed to chronic stress will either compensate or die. Even if they do compensate, their performance abilities will be depressed during the period of compensation. Also, even though fish can compensate, e.g., to a range of temperatures, they generally have a preferred temperature at which they perform best. Under less than ideal conditions, the fish is forced to allocate energy to compensatory functions. Also, under conditions of stress, basic metabolic functions have priority over such functions as growth and reproduction. Fish that show no growth, are slow to grow or will not reproduce can often be turned around by reducing stressors and providing conditions closer to ideal.
Stress alleviation
The closer an environment comes to ideal, the less undesirable stress is placed on fish in captivity. Providing good, stable water quality is probably the single most important factor in producing a positive environment. Elimination of factors producing anxiety and/or fear will contribute to a favorable environment. Good nutrition and appropriate exercise round out the major factors influencing good health.
There are several ways to relieve the stress of capture and transport. This can be accomplished by reducing the awareness of the fish. Tranquillizing (with such things as MS222) or placing the fish in darkness can help reduce its awareness. These techniques are most helpful during transportation. Also, adding salts and/or cooling the transportation water are techniques used by hatcheries to reduce transportation mortalities. As a corollary in most instances, capturing fish in cooler water results in less stress. The down side is that fish stressed under cold conditions have less capacity to compensate.
Smell
Koi have an acute sense of smell and, combined with their sense of taste, are the primary senses in locating food in a natural environment (even over sight). If a substance dissolves in water, it has the potential to be detected by their ‘nose.’
The olfactory organs (used to smell) are located at the base of the nostrils (called nares). Water does not flow to any other part of the body from the nares. They are exclusively used for olfaction (smelling), are paired and located between the eyes and the mouth. They are shaped like, and can be thought of as, small U-tubes into which water enters thru the leading or forward hole and exits thru the rear port or opening. Just behind the forward opening there is a flap of skin that directs water into the forward opening of the nare as the koi moves forward in the water. The movement of substances through the nares is aided by diffusion and by the motion of small hairs-like structures (cilia) within the nares.
Taste
Taste buds are numerous in and around the lips, mouth and tips of the barbels. Koi have two pairs of barbels. Three hundred years ago they had three pairs.

Lateral Line System
The lateral line is used to sense low frequency vibrations within the water and, since it is unlike any sense we poses, is difficult to imagine its perception by the fish. It has been described as “far touch” in that it allows fish to sense waves reflected from other objects and thus allows them to sense them at a distance. It is not like sonar but similar in that it does use low frequency waves, 1 to 200 Hz.
Hearing
Koi, like all fish, have no external ears. However, they can and do hear. This is accomplished by sensing vibrations in the water via the Weberian Ossicles, a group of bones that are connected on one end to the forward swim bladder and on the other end to the auditory center, a sensing organ that resembles our inner ear. It is believed that the sound is amplified as it emanates from the swim bladder through Weberian apparatus to the auditory center.
Acknowledgements
This section was made better by the review and comments of Sandy Yosha, DVM, Duncan Griffiths and Richard E. Carlson.
References:
Brewster, Bernice – “The big guide to koi digestion,” Koi Ponds and Gardens, Feb. 2002
Evans, David H. – The Physiology of Fishes, Second Edition; CRC Press.
Gratzek, John – Aquariology, Master Volume; Tetra Press.
Hulse, David – “It’s in the blood,” Koi Carp, Nov. 2000
Hulse, David – “The liver,” Koi Carp
Maslin, Paul - web page at: http://www.csuchico.edu/~pmaslin/ichthy/fshrsp.html
Reynolds, Paula – “Bouyancy aid,” Koi Carp, Feb. 2002
Saint-Erne,
Nicholas – “Advanced Koi Care” – unpublished.
Stoskopf, Michael – Fish Medicine; W.B. Saunders Company